 Tweet
Tweet
[Source: Eurosurveillance, full text: (LINK). Edited.]
Eurosurveillance, Volume 16, Issue 50, 15 December 2011
Rapid communications
S-OtrH3N2 viruses: use of sequence data for description of the molecular characteristics of the viruses and their relatedness to previously circulating H3N2 human viruses
B Lina ( )<SUP>1</SUP><SUP>,2</SUP>, M Bouscambert<SUP>1</SUP>, V Enouf<SUP>3</SUP>, D Rousset<SUP>3</SUP>, M Valette<SUP>1</SUP>, S van der Werf<SUP>3</SUP><SUP>,4</SUP><SUP>,5</SUP><SUP>,6</SUP>
)<SUP>1</SUP><SUP>,2</SUP>, M Bouscambert<SUP>1</SUP>, V Enouf<SUP>3</SUP>, D Rousset<SUP>3</SUP>, M Valette<SUP>1</SUP>, S van der Werf<SUP>3</SUP><SUP>,4</SUP><SUP>,5</SUP><SUP>,6</SUP>
Citation style for this article: Lina B, Bouscambert M, Enouf V, Rousset D, Valette M, van der Werf S. S-OtrH3N2 viruses: use of sequence data for description of the molecular characteristics of the viruses and their relatedness to previously circulating H3N2 human viruses. Euro Surveill. 2011;16(50):pii=20039. Available online: http://www.eurosurveillance.org/View...rticleId=20039
Date of submission: 02 December 2011 <HR>
Emergence of influenza viruses from the animal reservoir is a permanent challenge. The rapid description and immediate sharing of information on these viruses is invaluable for influenza surveillance networks and for pandemic preparedness. With the help of data generated from the United States Centers for Disease Control and Prevention, we provide here information on the swine?origin triple reassortant influenza A(H3N2) viruses detected in human cases in the north-east of the United States.
<HR>
On 23 November 2011, the United States (US) Centers for Disease Control and Prevention (CDC) reported three cases of documented infections with a triple reassortant influenza A(H3N2) virus of swine origin (S-OtrH3N2) that may have been acquired through human-to-human transmission [1]. In the last 10 years, at least 27 human cases of swine influenza virus infections had been observed in the US [2-4], all of which occurred after exposure to infected animals, and no human-to-human transmission had been reported so far. Noticeably, the 11 most recent cases recorded since August 2011 were due to viruses that had acquired the matrix (M) gene segment of the influenza A(H1N1)pdm09 virus through reassortment [1,5]. This may have resulted in enhanced transmission potential of the S-Otr H3N2 virus. Indeed, it has been reported that the acquisition by an S-Otr H1N1 virus of both the M and neuraminidase (NA) gene segments from the Eurasian swine lineage virus facilitated the emergence and the spread of the influenza A(H1N1)pdm09 virus [3]. In addition, epidemiological studies suggest that the four last cases reported between the 23 November and 9 December were observed in patients with no direct or indirect contact with swine, implying that limited human-to-human transmission has occurred.
This event raises concerns about the potential of such swine-origin viruses to establish a sustained human-to-human transmission and about our ability to fight against this virus, should it become pandemic. Sharing molecular data at a very early stage of emergence facilitates in silico analysis and risk assessment. Thanks to the CDC, the sequences of these viruses are available in the GISAID database (Table).
Table. GISAID reference numbers of S-Otr influenza A(H3N2) isolates from swine and humans in 2010 and 2011 used for the phylogenetic analysis

Phylogenetic relationship
Based on the data available, it was possible to draw a phylogenetic tree comprising haemagglutinin (HA) sequences from both human and S-Otr influenza A(H3N2) isolates, to determine if there were common characteristics between these two groups of viruses. We performed this analysis for a 966 nt sequence (nt 72 to 1,038) of the HA1 regions of the HA genes of all human reference strains used in the influenza vaccines between 1972 and 2011, of six S-Otr H3N2 viruses isolated from human cases in 2011, and of two S-Otr H3N2 viruses detected in the swine population in 2010 and 2011 (Figure 1).
Figure 1. Phylogenetic analysis of the haemagglutinin genes (nt 72-1,038) of 26 influenza A(H3N2) viruses (vaccine strains and S-Otr viruses)

This analysis shows that the human HA1 phylogenetically most closely related to the S-OtrH3N2 viruses was the A/Wuhan/359/95(H3N2) virus. This is consistent with the timing of introduction of the human H3N2 viruses into the swine population in North America [4]. This closest homology was confirmed when the evolutionary distances were computed using the Tamura-Nei method [6]. In our analysis, the 966 nt HA fragment of the S-Otr A/Iowa/08/2011(H3N2) virus showed 5.5% divergence from the A/Wuhan/359/95(H3N2), compared with 9.3% from the more recent A/Perth/16/2009(H3N2) virus. The evolutionary distances suggest a division of the human H3N2 viruses into two groups: one group of strains isolated between 1986 and 1999, which had the highest homology to S-Otr A/Iowa/08/2011(H3N2), and a second group comprising strains isolated before 1983 or after 1999, for which the divergence is larger than 8% and can reach as much as 11% (Figure 1).
In order to speculate on possible cross-protection, it is important to analyse differences in the antigenic sites. An alignment of the amino acid sequences of the HA1 subunit spanning the five antigenic sites of the HA protein [7] shows differences between the S-OtrH3N2 viruses and the human influenza A(H3N2) strains (Figure 2). Although we observed only few differences in the antigenic sites C, D and E, the differences in antigenic sites A and B were more significant.
Antigenic site B, closest to the receptor-binding site has been proposed to contribute most to the antigenic characteristics of the HA protein [8].
Figure 2. Amino acid sequence alignment of the haemagglutinin protein of 25 influenza A(H3N2) viruses (vaccine strains and S-Otr viruses) with antigenic sites A?E

Implications for diagnostics
The sequence analysis of the other gene segments like M, NP or NA also provided information on the capability of molecular diagnostic procedures to detect the S-Otr H3N2 virus. These viruses acquired the M gene segment from the human influenza A(H1N1)pdm09 virus. This implies that the generic detection through RT-PCR targeting the M gene will have very good sensitivity also for the S-Otr viruses. However, depending on possible mismatches in the primers and/or probes, the RT-PCR targeting the HA or NA gene segments may either be lacking sensitivity or possibly fail to detect the S-Otr H3N2 virus. Conversely, in terms of alertness and surveillance, the use of H3 and/or N2 subtyping RT-PCRs that would have equal sensitivity for seasonal human and swine-origin H3N2 viruses, should not be promoted at this stage because these procedures may fail to recognise cases of S-Otr H3N2 virus infection. Hence, for accurate detection and surveillance, specific RT-PCR methods should be developed, or alternatively, predefined algorithms with already existing discriminating molecular tools need to be implemented [9].
Lastly, the NA and M2 sequences available from the recent isolates suggest that, as for the influenza A(H1N1)pdm09 virus, antiviral drugs that block the M2 ion channel will not be effective because the M2 sequence carries the S31N mutation associated with resistance. No known genetic markers for resistance to NA inhibitors have been detected in these new strains so far. This should be confirmed by phenotypic assays.
Implications for immunological cross-protection
In case of the emergence of a zoonotic virus with an HA derived from previously circulating human viruses, it needs to be established whether or not infections with human influenza viruses in the past seasons or vaccinations confer cross-protection against the new viruses. Indeed, during the recent pandemic in 2009, it was observed that upon infection or vaccination of elderly people previously exposed to influenza A(H1N1) viruses that shared common epitopes with the emerging pandemic virus, efficient cross-protection was induced through memory immune cells [10]. The comparison of the five antigenic domains of past influenza A(H3N2) human viruses with those of the S-Otr viruses showed similarities and differences. Hence, it is impossible to predict if pre-existing immunity will be efficient against this virus, even if it seems likely that some cross-protection will exist; seroepidemiological surveys should be carried out to support or disprove this hypothesis. One must also keep in mind that if there is pre-existing immunity to this virus, it may occur in different age groups than observed with the influenza A(H1N1)pdm09 virus.
Conclusion
Overall, even if neither the evolution of these S-Otr H3N2 viruses nor their putative impact in the general population can be predicted enhanced surveillance with adapted diagnostic procedures will become necessary if these sporadic cases turn into sustained dissemination. According to the similarities observed between the sequences of the S-Otr and human H3 influenza viruses (especially those circulating before 1995), the likelihood of cross-protection is high, but should be confirmed with seroepidemiological studies. <HR>
Acknowledgements
This work was supported in part by the European Union Seventh Framework Programme [FP7/2007-2013] under the project PREDEMICS, EC [grant agreement number 278433].
<HR>
References-
------
Eurosurveillance, Volume 16, Issue 50, 15 December 2011
Rapid communications
S-OtrH3N2 viruses: use of sequence data for description of the molecular characteristics of the viruses and their relatedness to previously circulating H3N2 human viruses
B Lina (
- National Influenza Centre (Southern France), Hospices Civils de Lyon, Groupement Hospitalier Est, Bron, France
- Virpath, EA 4610, Faculty of Medicine R.T.H. Laennec, UCBL, Universit? de Lyon, Lyon, France
- Institut Pasteur, National Influenza Centre (Northern France), Paris, France
- Institut Pasteur, Unit of Molecular Genetics of RNA Viruses, Department of Virology, Paris France
- French National Centre for Scientific Research CNRS URA3015, Paris, France
- Universit? Paris Diderot, Sorbonne Paris Cit?, Unit of Molecular Genetics of RNA Viruses, Paris, France
Citation style for this article: Lina B, Bouscambert M, Enouf V, Rousset D, Valette M, van der Werf S. S-OtrH3N2 viruses: use of sequence data for description of the molecular characteristics of the viruses and their relatedness to previously circulating H3N2 human viruses. Euro Surveill. 2011;16(50):pii=20039. Available online: http://www.eurosurveillance.org/View...rticleId=20039
Date of submission: 02 December 2011 <HR>
Emergence of influenza viruses from the animal reservoir is a permanent challenge. The rapid description and immediate sharing of information on these viruses is invaluable for influenza surveillance networks and for pandemic preparedness. With the help of data generated from the United States Centers for Disease Control and Prevention, we provide here information on the swine?origin triple reassortant influenza A(H3N2) viruses detected in human cases in the north-east of the United States.
<HR>
On 23 November 2011, the United States (US) Centers for Disease Control and Prevention (CDC) reported three cases of documented infections with a triple reassortant influenza A(H3N2) virus of swine origin (S-OtrH3N2) that may have been acquired through human-to-human transmission [1]. In the last 10 years, at least 27 human cases of swine influenza virus infections had been observed in the US [2-4], all of which occurred after exposure to infected animals, and no human-to-human transmission had been reported so far. Noticeably, the 11 most recent cases recorded since August 2011 were due to viruses that had acquired the matrix (M) gene segment of the influenza A(H1N1)pdm09 virus through reassortment [1,5]. This may have resulted in enhanced transmission potential of the S-Otr H3N2 virus. Indeed, it has been reported that the acquisition by an S-Otr H1N1 virus of both the M and neuraminidase (NA) gene segments from the Eurasian swine lineage virus facilitated the emergence and the spread of the influenza A(H1N1)pdm09 virus [3]. In addition, epidemiological studies suggest that the four last cases reported between the 23 November and 9 December were observed in patients with no direct or indirect contact with swine, implying that limited human-to-human transmission has occurred.
This event raises concerns about the potential of such swine-origin viruses to establish a sustained human-to-human transmission and about our ability to fight against this virus, should it become pandemic. Sharing molecular data at a very early stage of emergence facilitates in silico analysis and risk assessment. Thanks to the CDC, the sequences of these viruses are available in the GISAID database (Table).
Table. GISAID reference numbers of S-Otr influenza A(H3N2) isolates from swine and humans in 2010 and 2011 used for the phylogenetic analysis

Phylogenetic relationship
Based on the data available, it was possible to draw a phylogenetic tree comprising haemagglutinin (HA) sequences from both human and S-Otr influenza A(H3N2) isolates, to determine if there were common characteristics between these two groups of viruses. We performed this analysis for a 966 nt sequence (nt 72 to 1,038) of the HA1 regions of the HA genes of all human reference strains used in the influenza vaccines between 1972 and 2011, of six S-Otr H3N2 viruses isolated from human cases in 2011, and of two S-Otr H3N2 viruses detected in the swine population in 2010 and 2011 (Figure 1).
Figure 1. Phylogenetic analysis of the haemagglutinin genes (nt 72-1,038) of 26 influenza A(H3N2) viruses (vaccine strains and S-Otr viruses)
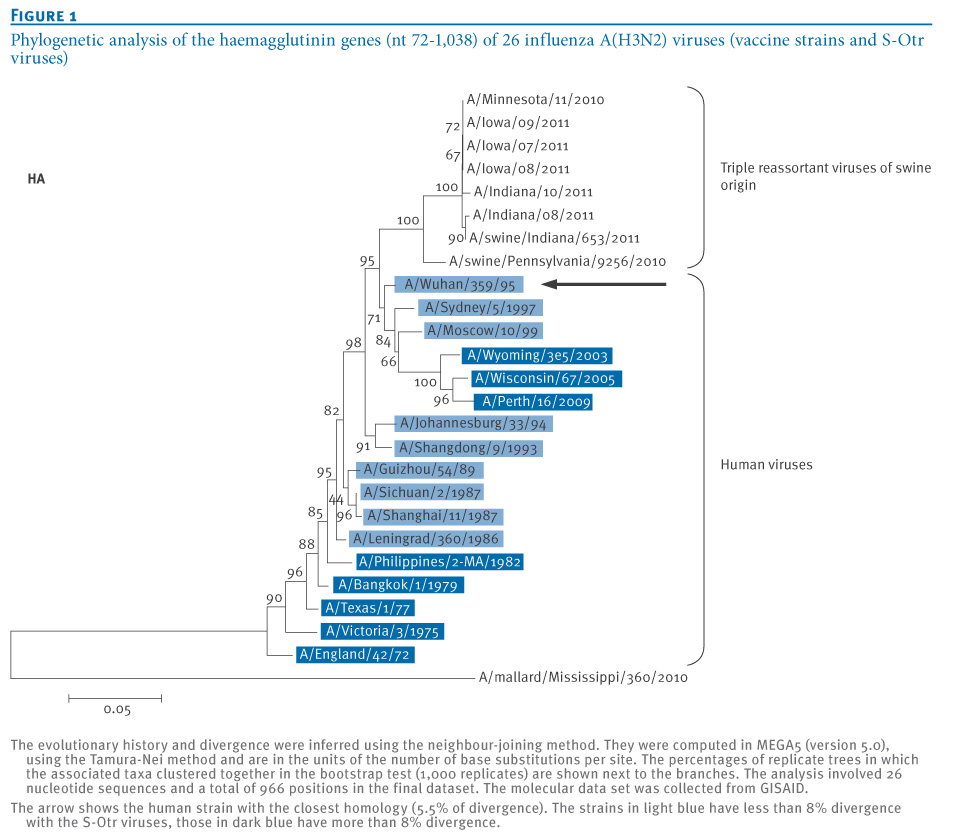
This analysis shows that the human HA1 phylogenetically most closely related to the S-OtrH3N2 viruses was the A/Wuhan/359/95(H3N2) virus. This is consistent with the timing of introduction of the human H3N2 viruses into the swine population in North America [4]. This closest homology was confirmed when the evolutionary distances were computed using the Tamura-Nei method [6]. In our analysis, the 966 nt HA fragment of the S-Otr A/Iowa/08/2011(H3N2) virus showed 5.5% divergence from the A/Wuhan/359/95(H3N2), compared with 9.3% from the more recent A/Perth/16/2009(H3N2) virus. The evolutionary distances suggest a division of the human H3N2 viruses into two groups: one group of strains isolated between 1986 and 1999, which had the highest homology to S-Otr A/Iowa/08/2011(H3N2), and a second group comprising strains isolated before 1983 or after 1999, for which the divergence is larger than 8% and can reach as much as 11% (Figure 1).
In order to speculate on possible cross-protection, it is important to analyse differences in the antigenic sites. An alignment of the amino acid sequences of the HA1 subunit spanning the five antigenic sites of the HA protein [7] shows differences between the S-OtrH3N2 viruses and the human influenza A(H3N2) strains (Figure 2). Although we observed only few differences in the antigenic sites C, D and E, the differences in antigenic sites A and B were more significant.
Antigenic site B, closest to the receptor-binding site has been proposed to contribute most to the antigenic characteristics of the HA protein [8].
Figure 2. Amino acid sequence alignment of the haemagglutinin protein of 25 influenza A(H3N2) viruses (vaccine strains and S-Otr viruses) with antigenic sites A?E

Implications for diagnostics
The sequence analysis of the other gene segments like M, NP or NA also provided information on the capability of molecular diagnostic procedures to detect the S-Otr H3N2 virus. These viruses acquired the M gene segment from the human influenza A(H1N1)pdm09 virus. This implies that the generic detection through RT-PCR targeting the M gene will have very good sensitivity also for the S-Otr viruses. However, depending on possible mismatches in the primers and/or probes, the RT-PCR targeting the HA or NA gene segments may either be lacking sensitivity or possibly fail to detect the S-Otr H3N2 virus. Conversely, in terms of alertness and surveillance, the use of H3 and/or N2 subtyping RT-PCRs that would have equal sensitivity for seasonal human and swine-origin H3N2 viruses, should not be promoted at this stage because these procedures may fail to recognise cases of S-Otr H3N2 virus infection. Hence, for accurate detection and surveillance, specific RT-PCR methods should be developed, or alternatively, predefined algorithms with already existing discriminating molecular tools need to be implemented [9].
Lastly, the NA and M2 sequences available from the recent isolates suggest that, as for the influenza A(H1N1)pdm09 virus, antiviral drugs that block the M2 ion channel will not be effective because the M2 sequence carries the S31N mutation associated with resistance. No known genetic markers for resistance to NA inhibitors have been detected in these new strains so far. This should be confirmed by phenotypic assays.
Implications for immunological cross-protection
In case of the emergence of a zoonotic virus with an HA derived from previously circulating human viruses, it needs to be established whether or not infections with human influenza viruses in the past seasons or vaccinations confer cross-protection against the new viruses. Indeed, during the recent pandemic in 2009, it was observed that upon infection or vaccination of elderly people previously exposed to influenza A(H1N1) viruses that shared common epitopes with the emerging pandemic virus, efficient cross-protection was induced through memory immune cells [10]. The comparison of the five antigenic domains of past influenza A(H3N2) human viruses with those of the S-Otr viruses showed similarities and differences. Hence, it is impossible to predict if pre-existing immunity will be efficient against this virus, even if it seems likely that some cross-protection will exist; seroepidemiological surveys should be carried out to support or disprove this hypothesis. One must also keep in mind that if there is pre-existing immunity to this virus, it may occur in different age groups than observed with the influenza A(H1N1)pdm09 virus.
Conclusion
Overall, even if neither the evolution of these S-Otr H3N2 viruses nor their putative impact in the general population can be predicted enhanced surveillance with adapted diagnostic procedures will become necessary if these sporadic cases turn into sustained dissemination. According to the similarities observed between the sequences of the S-Otr and human H3 influenza viruses (especially those circulating before 1995), the likelihood of cross-protection is high, but should be confirmed with seroepidemiological studies. <HR>
Acknowledgements
This work was supported in part by the European Union Seventh Framework Programme [FP7/2007-2013] under the project PREDEMICS, EC [grant agreement number 278433].
<HR>
References
- Centers for Disease Control and Prevention (CDC). Limited human-to-human transmission of novel influenza A (H3N2) virus - Iowa, November 2011. MMWR Morb Mortal Wkly Rep. 2011;60:1615-7.
- Shinde V, Bridges CB, Uyeki TM, Shu B, Balish A, Xu X, Lindstrom S, et al. Triple reassortant swine influenza A (H1) in humans in the United States, 2005--2009. N Engl J Med. 2009;360(25):2616-25.
- Brockwell-Staats C, Webster RG, Webby RJ. Diversity of influenza viruses in swine and the emergence of a novel human pandemic influenza A (H1N1). Influenza Other Respi Viruses. 2009;3(5):207-13.
- Shu B, Garten R, Emery S, Balish A, Cooper L, Sessions W, et al. Genetic analysis and antigenic characterization of swine origin influenza viruses isolated from humans in the United States, 1990-2010. Virology. 2012;422(1):151-60.
- Centers for Disease Control and Prevention (CDC). "Have You Heard?" CDC confirms two human infections with novel influenza viruses. Atlanta: CDC; 9 Dec 2011. Available from: http://www.cdc.gov/media/haveyouheard/stories/novel_influenza.html
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol Biol Evol. 2011;28(10):2731-9.
- Suwannakarn K, Chieochansin T, Thongmee C, Makkoch J, Praianantathavorn K, Theamboonlers A, et al. Molecular evolution of human H1N1 and H3N2 influenza A virus in Thailand, 2006-2009. PLoS One. 2010;5(3):e9717.
- Koel BF, Burke DF, Bestebroer TM, Van der Vliet S, Vervaet G, Skepner E, et al. 35 years of antigenic evolution of Influenza A/H3N2 virus is dictated by 7 aminoacid positions flanking the hemagglutinin receptor binding site. The fourth ESWI influenza conference; 11-14 Sept 2011; Malta.
- Sponseller BA, Strait E, Jergens A, Trujillo J, Harmon K, Koster L, et al. Influenza A Pandemic (H1N1) 2009 Virus Infection in Domestic Cat. Emerg Infect Dis. 2010;16(3):534-7.
- Miller E, Hoschler K, Hardelid P, Stanford E, Andrews N, Zambon M. Incidence of 2009 pandemic influenza A H1N1 infection in England: a cross-sectional serological study. Lancet. 2010;375(9720):1100-8.
------