 Tweet
Tweet
Re: China - H7N9 Human Isolates on Deposit at GISAID
The 211 amino residue is highly conserved in H7 Hemagglutinin as Isoleucine, but the position has seen previous variance to Valine in isolates from 7 horses almost 40 years ago and from 4 avian isolates prior to 1933. The Valine revision located on all emergent H7N9, including the Wet Market avian and environmental samples, is also recently seen as a polymorphism on Fatal Human H5N1. More importantly, this 211V (202V) that is Rare to H7 is conspicuously enough also Rare to H5. Although the Valine is found less than 150 times (147 Cases) in the GISAID database of 5,195 H5N1 HA segments, human fatalities are represented in 5 of the calendar years between 2004 and today, with 4 of those 6 deaths in a 14 month window from December 28, 2011 to February 9, 2013.
The polymorphism is known to be associated with Wet Markets, as 6 environmental samples from 2009 in Hebei and Guizhou traced to dominant Valine at this position within the H5N1 serotype. The 3 human H5N1 cases in the 2008/2009 period, including a fatality, are also associated with Wet Markets. One civet, one quail and one pigeon feature this polymorphism in H5N1. Additional avian host types numbering 18 and inclusive of commercial poultry species have carried the polymorphism in H5N1, H5N2 and H5N3 between 1993 and 2012. The change is not reported in avian H5N1 after 2011, but is reported in duck H5N2 and H5N3 during 2012.
Perhaps we are looking at the work of a potential host species transitioner for this H7 background . . . not quite mammalian, not quite avian . . . a buffer of some sort?
. . . . . . . . 211V [202V],
. . . . . . . . . . . . [H5N1 Human Rare, 13 Cases (Asia-12, Egypt-1)],
. . . . . . . . . . . . [H5N1 China Fatalities February 2013 (2)
. . . . . . . . . . . . . . . . 21F & 31M with host species transition signals,
. . . . . . . . . . . . . . . . . . . including 185S matching H7N9 wildtype],
. . . . . . . . . . . . [H5N1 China Fatality January 2012
. . . . . . . . . . . . . . . . 39M with host species transition signals,
. . . . . . . . . . . . . . . . . . . including 185S matching H7N9 wildtype],
. . . . . . . . . . . . [H5N1 China Fatality December 2011
. . . . . . . . . . . . . . . . 39M with host species transition signals],
. . . . . . . . . . . . [H5N1 China Human 2009
. . . . . . . . . . . . . . . . 23F & 29M Recovered, associated with Wet Market],
. . . . . . . . . . . . [H5N1 China Fatality December 2008
. . . . . . . . . . . . . . . . 19F, associated with Wet Market],
. . . . . . . . . . . . [H5N1 China Pigeon 2010],
. . . . . . . . . . . . [H5N1 China Wet Market Surveillance January 2009
. . . . . . . . . . . . . . . . Hebei & Guizhou from water, feces and environment],
. . . . . . . . . . . . [H5N1 Egypt Human 2011],
. . . . . . . . . . . . [H5N1 Vietnam Fatality January 2004 (1)],
. . . . . . . . . . . . [H5N1 Vietnam Human 2004 (4)],
. . . . . . . . . . . . [H5N1 Vietnam Civet 2005],
. . . . . . . . . . . . [H5N1 Vietnam Quail 2005],
. . . . . . . . . . . . [H5N2 Asia Avian],
. . . . . . . . . . . . [H5N2 Russia, Sweden, Switzerland, Portugal & Italy Avian],
. . . . . . . . . . . . [H5N3 Asia Avian],
. . . . . . . . . . . . [H5N3 Australia, Portugal & Italy Avian],
. . . . . . . . . . . . [H6N1],
. . . . . . . . . . . . [avH3N2 & pH1N1 wildtype],
. . . . . . . . . . . . [avH1N1farm: pH1N1 Ultimate Origin reservoir],
. . . . . . . . . . . . [WSN_1933, WSZ_1933],
. . . . . . . . . . . . [1918]
Potential H5N1 Involvement
in H7N9 Hemagglutinin Emergence
in H7N9 Hemagglutinin Emergence
The 211 amino residue is highly conserved in H7 Hemagglutinin as Isoleucine, but the position has seen previous variance to Valine in isolates from 7 horses almost 40 years ago and from 4 avian isolates prior to 1933. The Valine revision located on all emergent H7N9, including the Wet Market avian and environmental samples, is also recently seen as a polymorphism on Fatal Human H5N1. More importantly, this 211V (202V) that is Rare to H7 is conspicuously enough also Rare to H5. Although the Valine is found less than 150 times (147 Cases) in the GISAID database of 5,195 H5N1 HA segments, human fatalities are represented in 5 of the calendar years between 2004 and today, with 4 of those 6 deaths in a 14 month window from December 28, 2011 to February 9, 2013.
The polymorphism is known to be associated with Wet Markets, as 6 environmental samples from 2009 in Hebei and Guizhou traced to dominant Valine at this position within the H5N1 serotype. The 3 human H5N1 cases in the 2008/2009 period, including a fatality, are also associated with Wet Markets. One civet, one quail and one pigeon feature this polymorphism in H5N1. Additional avian host types numbering 18 and inclusive of commercial poultry species have carried the polymorphism in H5N1, H5N2 and H5N3 between 1993 and 2012. The change is not reported in avian H5N1 after 2011, but is reported in duck H5N2 and H5N3 during 2012.
Perhaps we are looking at the work of a potential host species transitioner for this H7 background . . . not quite mammalian, not quite avian . . . a buffer of some sort?
. . . . . . . . 211V [202V],
. . . . . . . . . . . . [H5N1 Human Rare, 13 Cases (Asia-12, Egypt-1)],
. . . . . . . . . . . . [H5N1 China Fatalities February 2013 (2)
. . . . . . . . . . . . . . . . 21F & 31M with host species transition signals,
. . . . . . . . . . . . . . . . . . . including 185S matching H7N9 wildtype],
. . . . . . . . . . . . [H5N1 China Fatality January 2012
. . . . . . . . . . . . . . . . 39M with host species transition signals,
. . . . . . . . . . . . . . . . . . . including 185S matching H7N9 wildtype],
. . . . . . . . . . . . [H5N1 China Fatality December 2011
. . . . . . . . . . . . . . . . 39M with host species transition signals],
. . . . . . . . . . . . [H5N1 China Human 2009
. . . . . . . . . . . . . . . . 23F & 29M Recovered, associated with Wet Market],
. . . . . . . . . . . . [H5N1 China Fatality December 2008
. . . . . . . . . . . . . . . . 19F, associated with Wet Market],
. . . . . . . . . . . . [H5N1 China Pigeon 2010],
. . . . . . . . . . . . [H5N1 China Wet Market Surveillance January 2009
. . . . . . . . . . . . . . . . Hebei & Guizhou from water, feces and environment],
. . . . . . . . . . . . [H5N1 Egypt Human 2011],
. . . . . . . . . . . . [H5N1 Vietnam Fatality January 2004 (1)],
. . . . . . . . . . . . [H5N1 Vietnam Human 2004 (4)],
. . . . . . . . . . . . [H5N1 Vietnam Civet 2005],
. . . . . . . . . . . . [H5N1 Vietnam Quail 2005],
. . . . . . . . . . . . [H5N2 Asia Avian],
. . . . . . . . . . . . [H5N2 Russia, Sweden, Switzerland, Portugal & Italy Avian],
. . . . . . . . . . . . [H5N3 Asia Avian],
. . . . . . . . . . . . [H5N3 Australia, Portugal & Italy Avian],
. . . . . . . . . . . . [H6N1],
. . . . . . . . . . . . [avH3N2 & pH1N1 wildtype],
. . . . . . . . . . . . [avH1N1farm: pH1N1 Ultimate Origin reservoir],
. . . . . . . . . . . . [WSN_1933, WSZ_1933],
. . . . . . . . . . . . [1918]
- Ongoing Sequence Analysis - Open-Access, Full-Text
- GISAID Citation
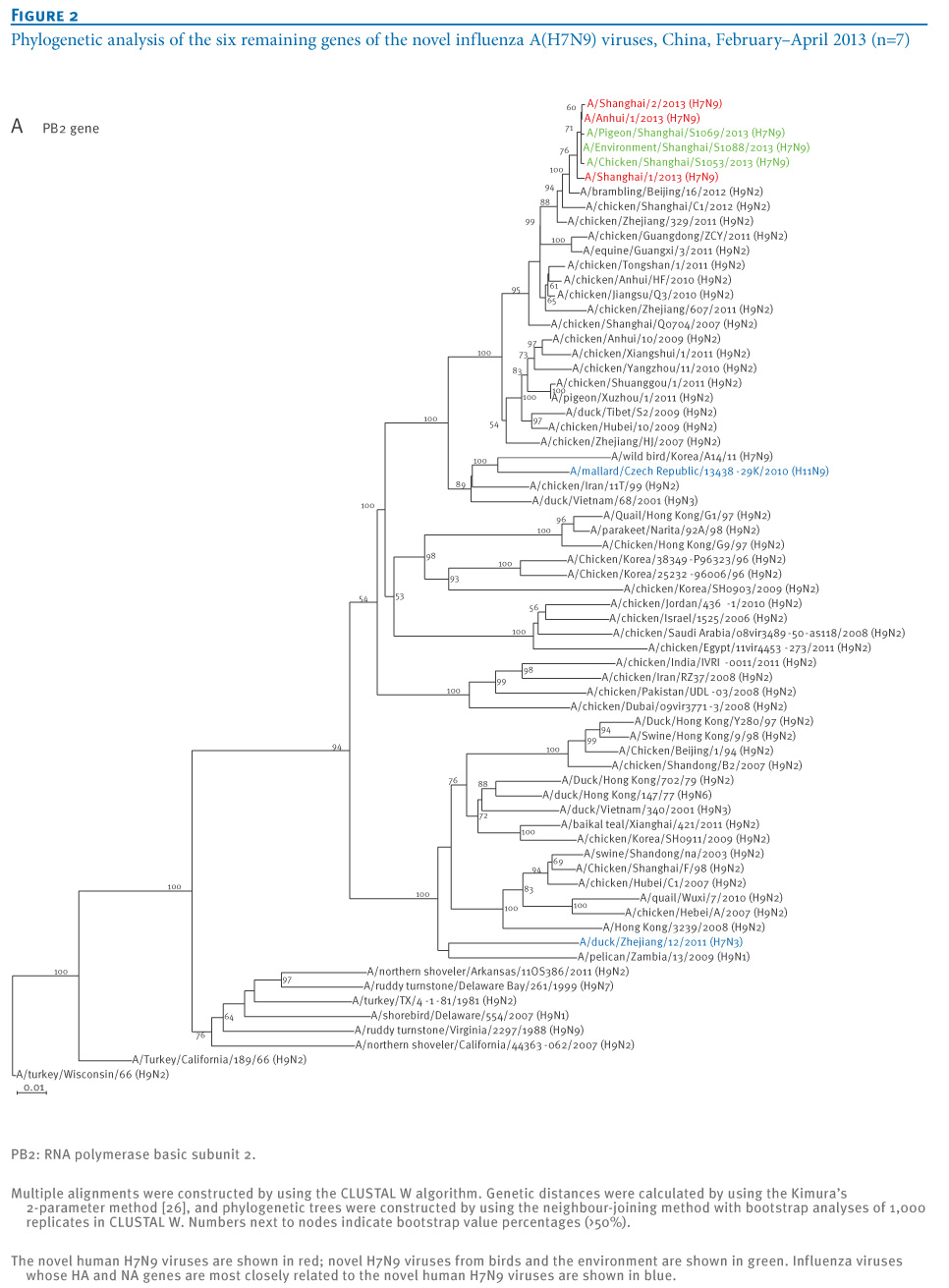
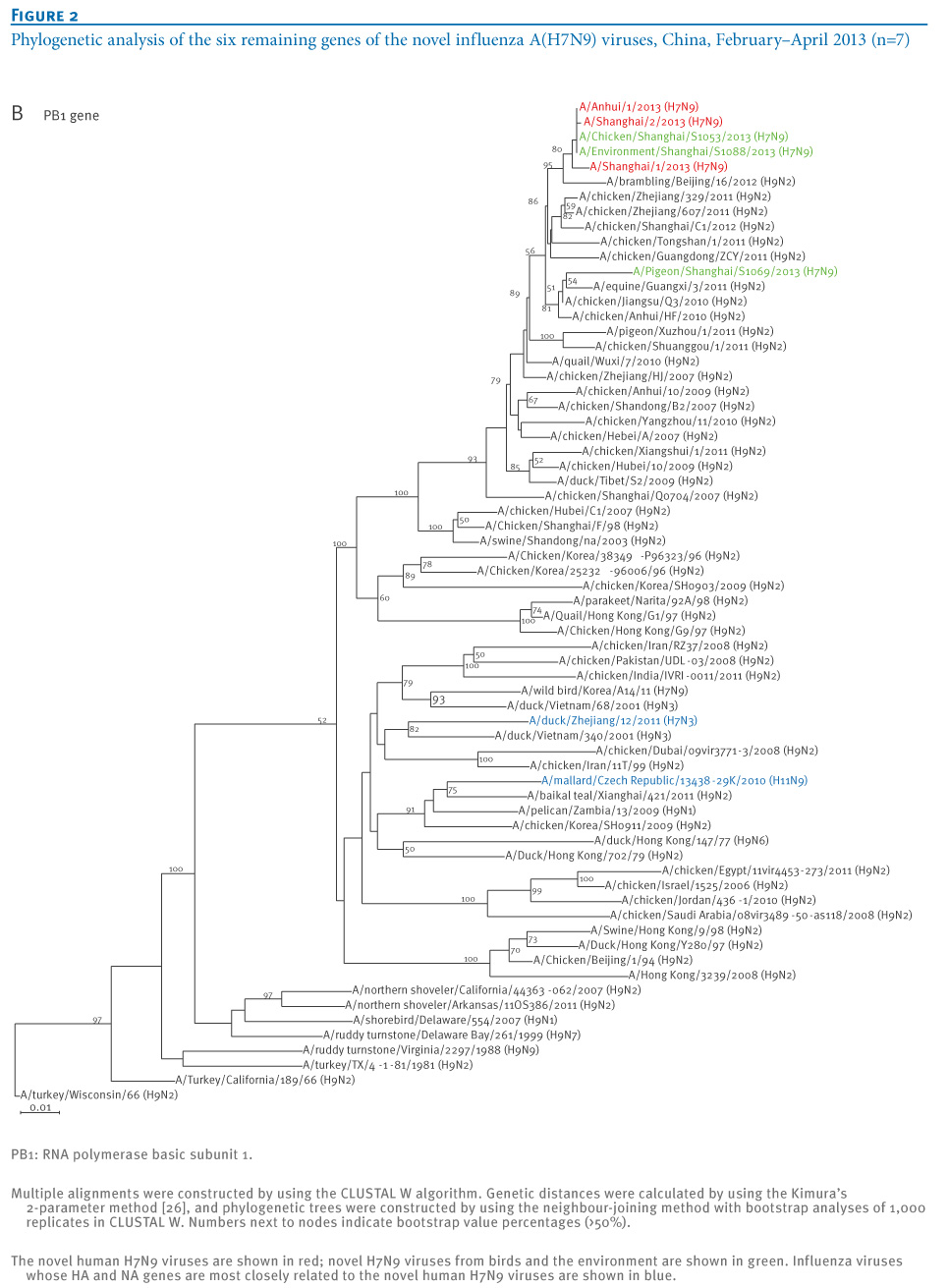
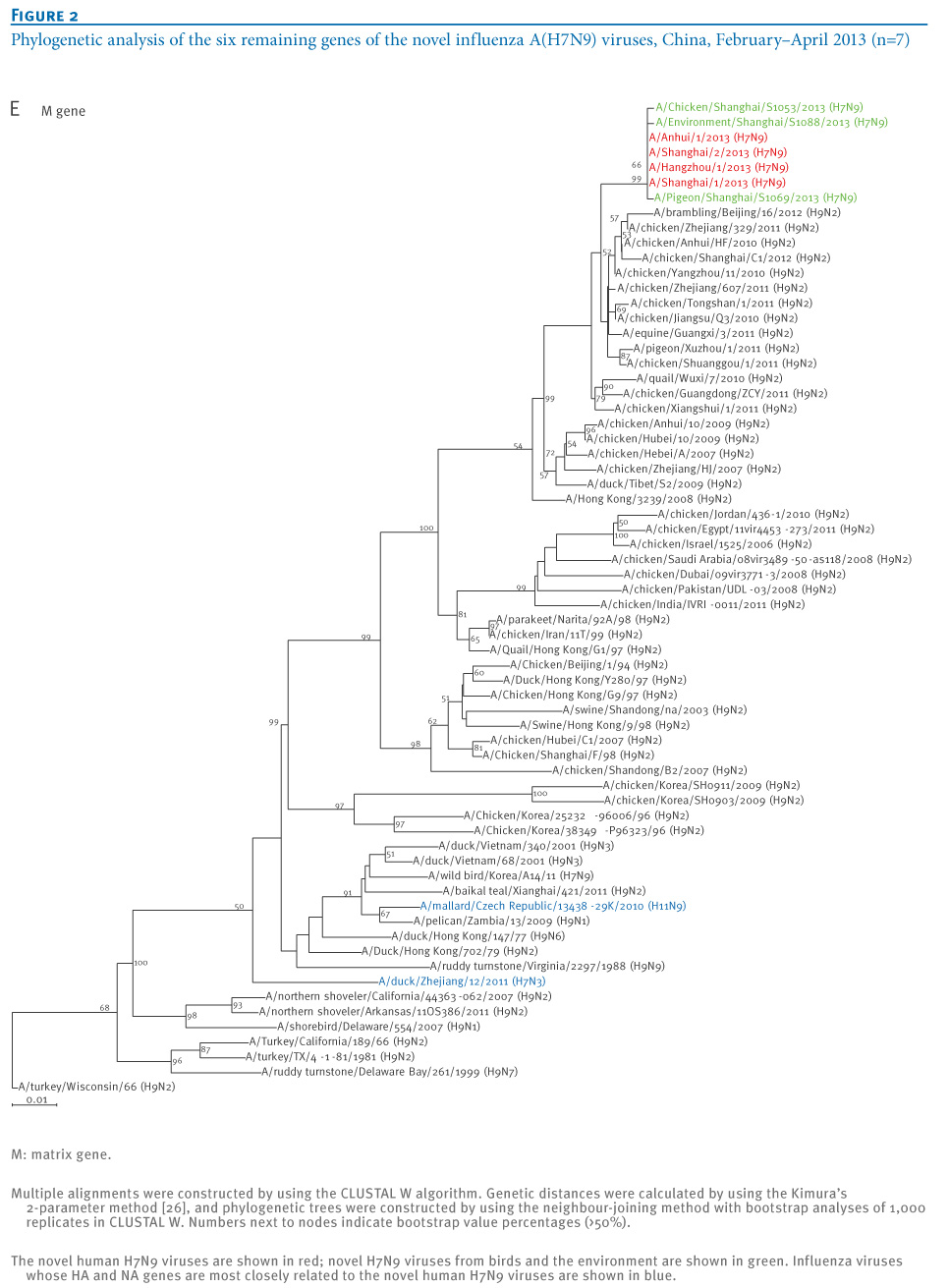
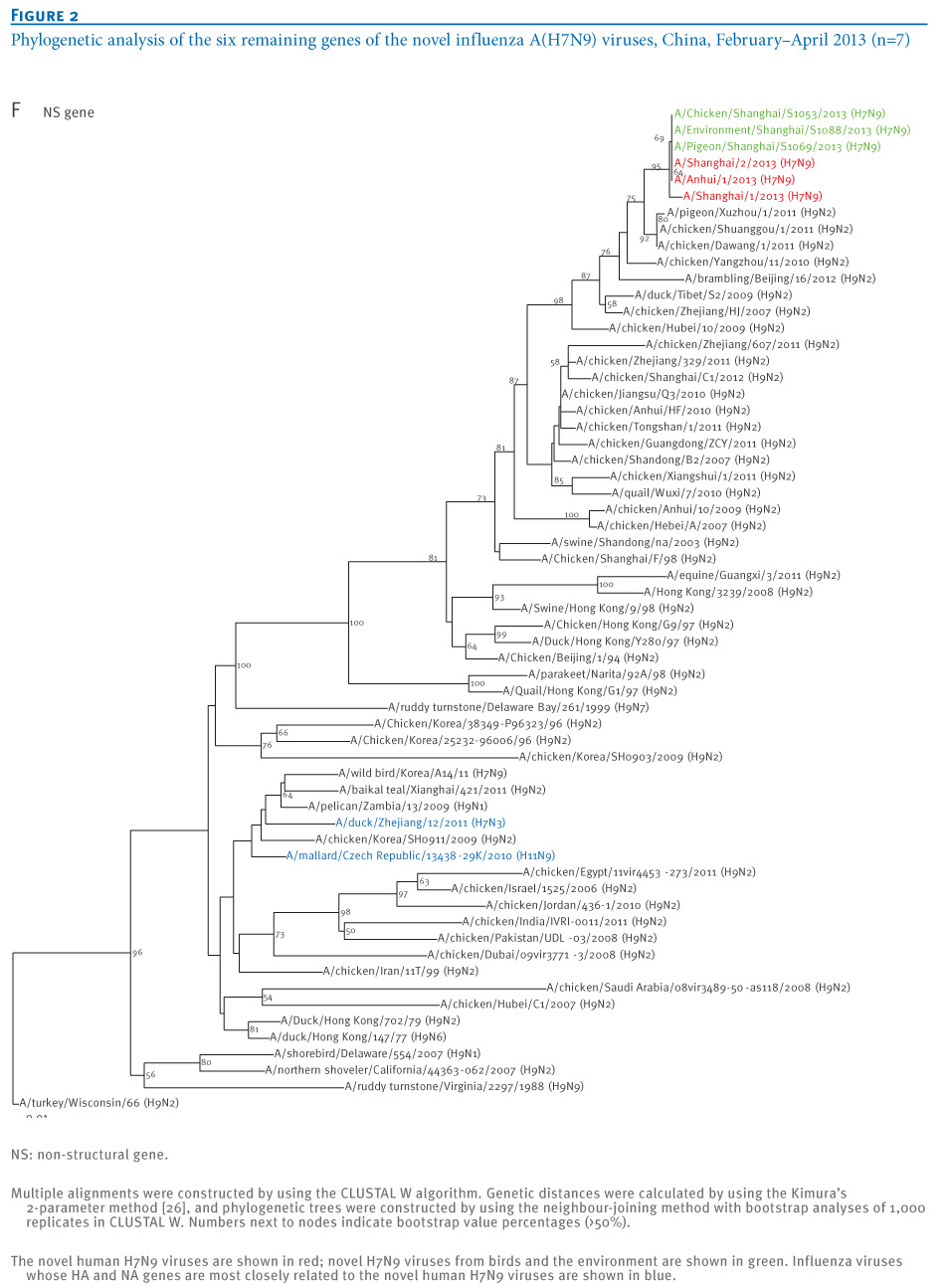
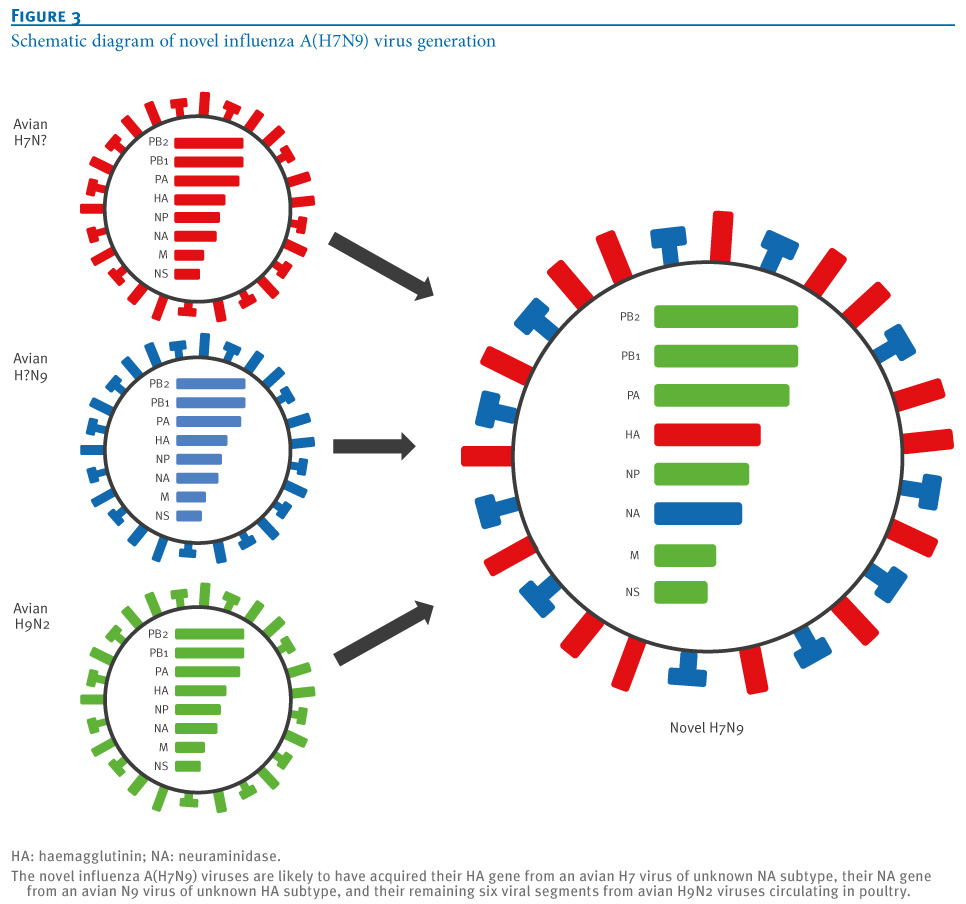

 oNotPromoteQF/> <w:LidThemeOther>EN-US</w:LidThemeOther> <w:LidThemeAsian>X-NONE</w:LidThemeAsian> <w:LidThemeComplexScript>X-NONE</w:LidThemeComplexScript> <w:Compatibility> <w:BreakWrappedTables/> <w:SnapToGridInCell/> <w:WrapTextWithPunct/> <w:UseAsianBreakRules/> <w
oNotPromoteQF/> <w:LidThemeOther>EN-US</w:LidThemeOther> <w:LidThemeAsian>X-NONE</w:LidThemeAsian> <w:LidThemeComplexScript>X-NONE</w:LidThemeComplexScript> <w:Compatibility> <w:BreakWrappedTables/> <w:SnapToGridInCell/> <w:WrapTextWithPunct/> <w:UseAsianBreakRules/> <w
Comment