 Tweet
Tweet
Eurosurveillance: Early Season Flu Surveillance & Vaccine Effectiveness (VE) - Canada
#13,114
Each year about this time we begin to get preliminary reports on the epidemiology, and vaccine effectiveness, of the winter flu season. With flu season likely only halfway over, things could change, and so we'll get a much better summation next summer.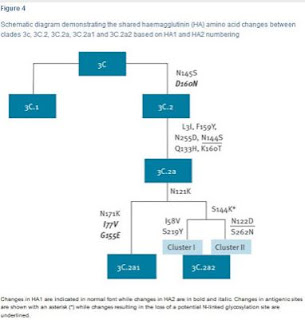
Not unexpectedly, the H3N2 component's early VE is much lower this year, at only 17%, but the influenza B component is doing much better at 55%, even though the trivalent shot often used in Canada contained a Victoria strain this year, suggesting some cross protection.
Some excerpts from a much longer, and far more detailed report. Follow the link to read it in its entirety. .
We will be seeing more of these types of preliminary assessments come out in the weeks ahead from all over the world. The flu epidemic in the United States has been far more H3N2-centric than what Canada has seen, and we're seeing influenza B, and even H1N1, dominating in other parts of the world this winter.
All of which means we're going to have a lot of diverse, and possibly conflicting, data to look over in the coming months.
http://afludiary.blogspot.com/2018/0...eason-flu.html
 |
| Canada Flu Watch Week 3 |
#13,114
Each year about this time we begin to get preliminary reports on the epidemiology, and vaccine effectiveness, of the winter flu season. With flu season likely only halfway over, things could change, and so we'll get a much better summation next summer.
Midway through last year's flu season, we saw a Eurosurveillance report Interim estimates of 2016/17 vaccine effectiveness against influenza A(H3N2), Canada, January 2017 by DM Skowronski et al. which found an adjusted VE of 42%.
But after reports this fall of only a 10% VE against H3N2 in Australia, the expectation for this winter was lowered (see ECDC:H3N2 Flu Vaccine Component Likely `Suboptimal'). Just before Christmas, the ECDC: Rapid Risk Assessment For Seasonal Flu EU/EEA 2017-18, stated the following:• Vaccine effectiveness against A(H1N1)pdm09 is expected to be high, whereas effectiveness against A(H3N2), the most prevalent type A virus, is likely to be low.
• B/Yamagata viruses are dominant so far, and vaccine effectiveness against this virus will be low as it is not included in the most widely used trivalent vaccine. Some effectiveness could be expected from cross-protection as a result of vaccination against B/Victoria virus.
Over the past few months we've looked repeatedly at some of the challenges posed by the H3N2 virus, and the problems of crafting effective vaccines against this particular subtype, including:Today Danuta M Skowronski et al. are back with an early look at Canada's H3N2/Influenza B dominated flu season, and at this year's vaccine performance.A bit unexpectedly, the vast majority of H3N2 viruses analyzed were from a single subgroup of clade 3C.2a with T131K + R142K + R261Q substitutions, as opposed to last season when 80% belonged to clade 3C.2a1.
The 3C.2a clade emerged in 2014, and had been the dominate clade until last winter when it further split into 3C.2a1 and 3C.2a2 (see Eurosurveillance: Emergence Of A Novel Subclade Of Seasonal A/H3N2 - London). The following Eurosurveillance chart shows the recent evolution of H3N2 clade 3C. Not unexpectedly, the H3N2 component's early VE is much lower this year, at only 17%, but the influenza B component is doing much better at 55%, even though the trivalent shot often used in Canada contained a Victoria strain this year, suggesting some cross protection.
Some excerpts from a much longer, and far more detailed report. Follow the link to read it in its entirety. .
Rapid communication Open Access
 Early season co-circulation of influenza A(H3N2) and B(Yamagata): interim estimates of 2017/18 vaccine effectiveness, Canada, January 2018
Early season co-circulation of influenza A(H3N2) and B(Yamagata): interim estimates of 2017/18 vaccine effectiveness, Canada, January 2018
Danuta M Skowronski1,2, Catharine Chambers1, Gaston De Serres3,4,5, James A Dickinson6, Anne-Luise Winter7, Rebecca Hickman1, Tracy Chan1, Agatha N Jassem1,2, Steven J Drews8,9, Hugues Charest3, Jonathan B Gubbay7,10, Nathalie Bastien11, Yan Li11, Mel Krajden1,2
The 2017/18 influenza season in Canada has been characterised by co-circulation of influenza A(H3N2) and B(Yamagata) viruses, the latter unusual so early in the season [1]. Most European countries are also experiencing simultaneous influenza A and B epidemics, with B(Yamagata) predominating [2], whereas the United States (US) has experienced a substantial epidemic due predominantly to influenza A(H3N2) [3].
The 2017/18 trivalent influenza vaccine (TIV) includes influenza A/Hong Kong/4801/2014(H3N2)-like (clade 3C.2a) and B/Brisbane/60/2008(Victoria-lineage)-like (clade 1A) antigens. The quadrivalent influenza vaccine (QIV) contains an additional influenza B/Phuket/3073/2013(Yamagata-lineage)-like (clade 3) antigen. The same components were included in the 2016/17 northern and 2017 southern hemisphere vaccines [4]. Low vaccine effectiveness (VE) for the 2017/18 season has been anticipated following the interim report from Australia indicating VE of just 10% during its 2017 influenza A(H3N2) epidemic [5]. In the context of exclusive QIV use, Australia reported higher VE of 57% against co-circulating influenza B viruses [5]. Here we report interim 2017/18 VE estimates for influenza A(H3N2) and influenza B from participating provinces of the Canadian Sentinel Practitioner Surveillance Network (SPSN), where QIV comprised less than one third of vaccine doses distributed overall through the publicly funded campaign.
(SNIP)
Most sequenced influenza A(H3N2) viruses belonged to genetic clade 3C.2a (213/229; 93%) and of these most (204/213; 96%) belonged to a single genetic subgroup of 3C.2a (denoted subgroup 3 by nextflu.org [10]), bearing antigenic site A substitutions T131K and R142K and antigenic site E substitution R261Q (Table 1). Overall 89% of influenza A(H3N2) viruses belonged to clade 3C.2a subgroup 3, which is similar to other surveillance observations from Canada (83%) (Figure 2) and to recent reports from Europe [11]. However, this profile for the 2017/18 season is different from that found by the Canadian SPSN during 2016/17 or by Australia during its 2017 epidemic, when a greater mix of genetic variants contributed to interim analyses and only 14% and 7%, respectively, of influenza A(H3N2) viruses belonged to subgroup 3 (Figure 2).

(SNIP)Danuta M Skowronski1,2, Catharine Chambers1, Gaston De Serres3,4,5, James A Dickinson6, Anne-Luise Winter7, Rebecca Hickman1, Tracy Chan1, Agatha N Jassem1,2, Steven J Drews8,9, Hugues Charest3, Jonathan B Gubbay7,10, Nathalie Bastien11, Yan Li11, Mel Krajden1,2
The 2017/18 influenza season in Canada has been characterised by co-circulation of influenza A(H3N2) and B(Yamagata) viruses, the latter unusual so early in the season [1]. Most European countries are also experiencing simultaneous influenza A and B epidemics, with B(Yamagata) predominating [2], whereas the United States (US) has experienced a substantial epidemic due predominantly to influenza A(H3N2) [3].
The 2017/18 trivalent influenza vaccine (TIV) includes influenza A/Hong Kong/4801/2014(H3N2)-like (clade 3C.2a) and B/Brisbane/60/2008(Victoria-lineage)-like (clade 1A) antigens. The quadrivalent influenza vaccine (QIV) contains an additional influenza B/Phuket/3073/2013(Yamagata-lineage)-like (clade 3) antigen. The same components were included in the 2016/17 northern and 2017 southern hemisphere vaccines [4]. Low vaccine effectiveness (VE) for the 2017/18 season has been anticipated following the interim report from Australia indicating VE of just 10% during its 2017 influenza A(H3N2) epidemic [5]. In the context of exclusive QIV use, Australia reported higher VE of 57% against co-circulating influenza B viruses [5]. Here we report interim 2017/18 VE estimates for influenza A(H3N2) and influenza B from participating provinces of the Canadian Sentinel Practitioner Surveillance Network (SPSN), where QIV comprised less than one third of vaccine doses distributed overall through the publicly funded campaign.
(SNIP)
Most sequenced influenza A(H3N2) viruses belonged to genetic clade 3C.2a (213/229; 93%) and of these most (204/213; 96%) belonged to a single genetic subgroup of 3C.2a (denoted subgroup 3 by nextflu.org [10]), bearing antigenic site A substitutions T131K and R142K and antigenic site E substitution R261Q (Table 1). Overall 89% of influenza A(H3N2) viruses belonged to clade 3C.2a subgroup 3, which is similar to other surveillance observations from Canada (83%) (Figure 2) and to recent reports from Europe [11]. However, this profile for the 2017/18 season is different from that found by the Canadian SPSN during 2016/17 or by Australia during its 2017 epidemic, when a greater mix of genetic variants contributed to interim analyses and only 14% and 7%, respectively, of influenza A(H3N2) viruses belonged to subgroup 3 (Figure 2).

Discussion
In most other interim analyses by the Canadian SPSN, type B viruses comprised less than 10% of influenza detections, whereas in 2017/18, they were identified in an equal proportion with influenza A(H3N2) [7-9]. Although the reasons for an earlier influenza B onset are unclear, Canada experienced a substantial influenza A(H3N2) epidemic in 2016/17 that may have altered population immunity and the overall 2017/18 influenza A(H3N2) contribution [9].
Nearly all (93%) characterised influenza A(H3N2)viruses were clade 3C.2a, a change from 2016/17 when most (80%) of the A(H3N2) viruses instead belonged to clade 3C.2a1 [9]. Furthermore, a single subgroup of clade 3C.2a with T131K + R142K + R261Q substitutions (i.e. nextflu subgroup 3 [10]) is currently predominating (89% of influenza A(H3N2) viruses), whereas a more heterogeneous mix of genetic variants contributed in Canada during 2016/17 [9] and in Australia during their 2017 epidemic [5]. Changes in the proportionate contribution and emerging predominance of clade 3C.2a variants among circulating influenza A(H3N2) viruses are important to monitor globally. The World Health Organization will decide in February 2018 whether to update the current clade 3C.2a vaccine antigen for the 2018/19 northern hemisphere vaccine, having already chosen a clade 3C.2a1 strain for the southern hemisphere’s 2018 vaccine [4].
In most other interim analyses by the Canadian SPSN, type B viruses comprised less than 10% of influenza detections, whereas in 2017/18, they were identified in an equal proportion with influenza A(H3N2) [7-9]. Although the reasons for an earlier influenza B onset are unclear, Canada experienced a substantial influenza A(H3N2) epidemic in 2016/17 that may have altered population immunity and the overall 2017/18 influenza A(H3N2) contribution [9].
Nearly all (93%) characterised influenza A(H3N2)viruses were clade 3C.2a, a change from 2016/17 when most (80%) of the A(H3N2) viruses instead belonged to clade 3C.2a1 [9]. Furthermore, a single subgroup of clade 3C.2a with T131K + R142K + R261Q substitutions (i.e. nextflu subgroup 3 [10]) is currently predominating (89% of influenza A(H3N2) viruses), whereas a more heterogeneous mix of genetic variants contributed in Canada during 2016/17 [9] and in Australia during their 2017 epidemic [5]. Changes in the proportionate contribution and emerging predominance of clade 3C.2a variants among circulating influenza A(H3N2) viruses are important to monitor globally. The World Health Organization will decide in February 2018 whether to update the current clade 3C.2a vaccine antigen for the 2018/19 northern hemisphere vaccine, having already chosen a clade 3C.2a1 strain for the southern hemisphere’s 2018 vaccine [4].
Our 2017/18 interim VE estimate of 17% (95% CI: −14 to 40) is less than half that reported for the same A(H3N2) vaccine in 2016/17, including interim analyses by the Canadian SPSN (42%; 95% CI: 18 to 59) [9], the US Flu VE Network (43%; 95% CI: 29 to 54) [12] and the European I-MOVE Network (38%; 95% CI: 21 to 51) [13]. Our estimate is also lower than end-of-season estimates from Canada (37%; 95% CI: 20 to 51) [14] and the US (34%; 95% CI: 24 to 42) for 2016/17 [15], and lower than is expected generally for influenza A(H3N2) vaccines (33%; 95% CI: 26 to 39) [16].
Our 2017/18 interim VE for influenza A(H3N2) is more comparable to the 2017 southern hemisphere interim VE of 10% (95% CI: −16 to 31) reported from Australia [5]. Differences in virological and participant profiles, as well as the stage of the epidemic, have to be taken into account when comparing VE estimates across studies. Working-age adults comprised the majority of participants in both studies and the 2017/18 interim VE against influenza A(H3N2) among Canadian SPSN participants 20–64-years-old (10%; 95% CI: −31 to 39) is also comparable to the 2017 estimate reported from Australia for 15–64-year-olds (16%; 95% CI: −11 to 36). Sample size for other age groups (e.g. children, elderly adults) was too limited to derive reliable interim estimates or to inform protection in specific high-risk groups.
Our 2017/18 interim VE for influenza A(H3N2) is more comparable to the 2017 southern hemisphere interim VE of 10% (95% CI: −16 to 31) reported from Australia [5]. Differences in virological and participant profiles, as well as the stage of the epidemic, have to be taken into account when comparing VE estimates across studies. Working-age adults comprised the majority of participants in both studies and the 2017/18 interim VE against influenza A(H3N2) among Canadian SPSN participants 20–64-years-old (10%; 95% CI: −31 to 39) is also comparable to the 2017 estimate reported from Australia for 15–64-year-olds (16%; 95% CI: −11 to 36). Sample size for other age groups (e.g. children, elderly adults) was too limited to derive reliable interim estimates or to inform protection in specific high-risk groups.
All influenza vaccine manufacturing in Canada is egg-based. Mutations that arise from egg adaptation of the vaccine strain may affect VE, an issue also identified for the current season’s A(H3N2) vaccine component [17,18]. In Canada this season, antigenic characterisation of influenza A(H3N2) viruses has only been presented in relation to a cell-propagated version of the vaccine reference strain; characterisation against an egg-based version has not been reported [1]. Among the small subset of Canadian viruses that could be successfully characterised, all were considered antigenically similar to the cell-propagated vaccine strain [1]. Conversely, where relatedness to the egg-propagated version of the vaccine strain has been specifically explored elsewhere, more variability has been identified, with a greater proportion of viruses considered antigenically distinct from the egg-propagated version [5,11,19].
We found higher VE of 55% (95% CI: 38 to 68) against influenza B despite prominent use of TIV containing a B(Victoria) antigen that was lineage-mismatched to almost exclusively B(Yamagata) viruses. Approximately 70% of vaccine doses distributed in SPSN provinces during the 2017/18 season were TIV, albeit with regional variation that will be explored in end-of-season analyses. Substantial cross-lineage VE for influenza B has been observed previously [20], including during the prior 2016/17 season in Canada when VE against lineage-mismatched influenza B using the same B(Victoria) TIV component was 73% (95% CI: 52 to 84) [14] and QIV comprised an even smaller proportion (< 25%) of vaccine doses distributed. Our estimate for the current season is comparable to the interim VE of 57% (95% CI: 41 to 69) for influenza B reported from Australia, despite exclusive use of QIV in that country [5].
Other agent–host and immuno–epidemiological interactions, including birth cohort effects induced by differential prime–boost exposures, may also play a role in VE [21]. The effect of prior vaccination history was not assessed here owing to sample size limitations, but will be explored as part of the end-of-season analyses.
Conclusions
As reported from Australia for the 2017 southern hemisphere vaccine, interim estimates from Canada for the 2017/18 northern hemisphere vaccine indicate low VE of less than 20% against influenza A(H3N2), notably among working-age adults. While the influenza A(H3N2) epidemic continues, adjunct protective measures should be reinforced to minimise the associated disease burden in high-risk individuals [22]. Interim 2017/18 VE estimates against influenza B are higher at 55% despite prominent TIV use, suggesting cross-lineage protection.
(Continue .. . )We found higher VE of 55% (95% CI: 38 to 68) against influenza B despite prominent use of TIV containing a B(Victoria) antigen that was lineage-mismatched to almost exclusively B(Yamagata) viruses. Approximately 70% of vaccine doses distributed in SPSN provinces during the 2017/18 season were TIV, albeit with regional variation that will be explored in end-of-season analyses. Substantial cross-lineage VE for influenza B has been observed previously [20], including during the prior 2016/17 season in Canada when VE against lineage-mismatched influenza B using the same B(Victoria) TIV component was 73% (95% CI: 52 to 84) [14] and QIV comprised an even smaller proportion (< 25%) of vaccine doses distributed. Our estimate for the current season is comparable to the interim VE of 57% (95% CI: 41 to 69) for influenza B reported from Australia, despite exclusive use of QIV in that country [5].
Other agent–host and immuno–epidemiological interactions, including birth cohort effects induced by differential prime–boost exposures, may also play a role in VE [21]. The effect of prior vaccination history was not assessed here owing to sample size limitations, but will be explored as part of the end-of-season analyses.
Conclusions
As reported from Australia for the 2017 southern hemisphere vaccine, interim estimates from Canada for the 2017/18 northern hemisphere vaccine indicate low VE of less than 20% against influenza A(H3N2), notably among working-age adults. While the influenza A(H3N2) epidemic continues, adjunct protective measures should be reinforced to minimise the associated disease burden in high-risk individuals [22]. Interim 2017/18 VE estimates against influenza B are higher at 55% despite prominent TIV use, suggesting cross-lineage protection.
We will be seeing more of these types of preliminary assessments come out in the weeks ahead from all over the world. The flu epidemic in the United States has been far more H3N2-centric than what Canada has seen, and we're seeing influenza B, and even H1N1, dominating in other parts of the world this winter.
All of which means we're going to have a lot of diverse, and possibly conflicting, data to look over in the coming months.
http://afludiary.blogspot.com/2018/0...eason-flu.html